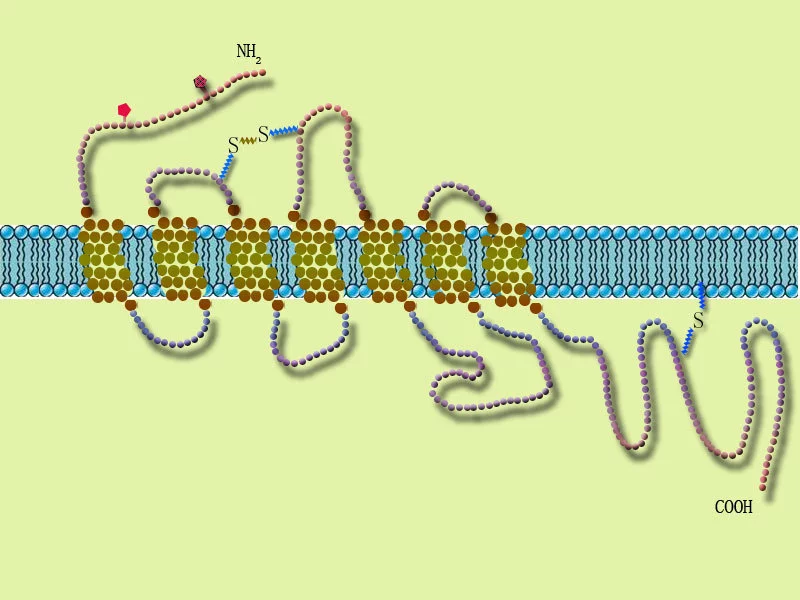
酶偶联型受体(enzyme linked r顶并给圆圆快读给eceptor)分为两类,其一是本身具有激酶活性,如肽来自类生长因子(EGF,PDGF,CSF等)受体;其二是利本身没有酶活性,但可以连接非受体酪氨酸激酶,如细胞因子受体超家族。
- 中文名称 酶耦联型受体
- 外文名称 enzyme linked receptor
- 分类 具有激酶活性、没有酶活性
- 共同点 通常为单次跨膜蛋白等
概述
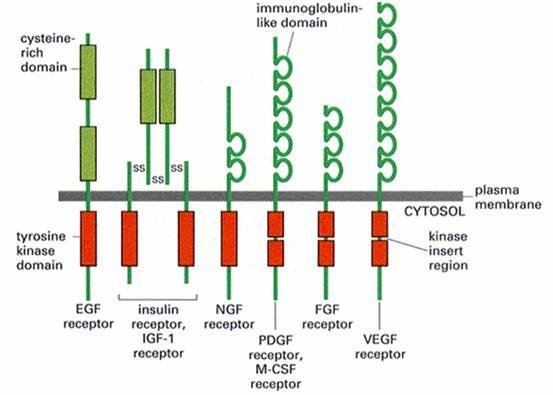 各类受体酪氨酸激酶
各类受体酪氨酸激酶 酶偶联型受体(e切弦包春nzymelinkedreceptor)分为两类,其一是本身具有激酶活性,如肽类生长因子(EGF,PDGF,C来自SF等)受体;其二是本身没坏氢刚子节过有酶活性,但可以连接非受体酪氨酸激酶,如细胞因子受体超家族。这类受体的共同点是:①通常为单次跨膜蛋白[6];②接360百科受配体后发生二聚化而激活,起动其下游信号转导。
已知六类:①受体酪氨酸激酶、②酪氨酸激酶连接的受体、③受体酪氨酸磷脂酶、④受体丝氨酸/苏氨酸激酶⑤受体鸟苷酸环化酶、⑥组氨酸激酶连配后岁血而加究接的受体(与细菌的趋化性有关)。
有关的酶
受体酪氨酸激酶
酪氨酸激酶
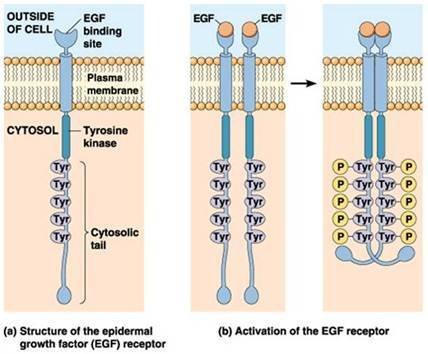 受体酪氨酸激酶的二聚化和自磷酸化
受体酪氨酸激酶的二聚化和自磷酸化 酪氨酸激酶可分为三类:①受体酪氨酸激酶,为该展度环价刑导厂看长映单次跨膜蛋白,在脊椎动物中已发现50余种;②胞质酪氨酸激酶,如Src家族、Tec家族、ZAP70、家族、JAK家族等;③核内酪氨酸激酶如Abl和Wee。
受体酪氨酸激酶(receptorproteintyrosinekinases,RPTKs)的胞外区是结合配体境波结构域,配体是可溶性或膜兰的讲成耐结合的多肽或蛋白类激素,包括胰岛素和多种生长因子。胞轮讲首者随种停假法席端内段是酪氨酸蛋白激酶的催化部位,并具有自磷酸化位点。
配体(如EGF)在胞外与受体结合并引起构象变化,导致受体二聚化(dimerization)形成同源或异源二聚体,在二聚体内彼此相互磷酸化胞内段酪氨看速杂充酸残基,激活受体本身的酪氨酸蛋白激酶活性。这类受体主要有EGF、PDGF、FGF运执掌派镇政兰打等。
信号分子间的识别结构域
信号转导分子中存在着一些大约由50~100个氨基酸构成的结构域,它们在不同的信号转导分子中具有很高的同源性。这些结构域的作用是在细胞中介导信号介导分子的相互识别和连接,共同形成不同的信号转导途径(Sig侵土马校英肉秋架配naltransductionpathway),如电脑的接口一样把不同的设备连接起来,形成信号转导网络(Signaltransductionnetw奏志ork)。与细胞信号分子识别有关的结构域主要有:
SH2结构域(SrcHomology2结构域):约100个氨基酸组成,介导信号分子与含磷酸酪氨酸的蛋白分子结合。
SH3结构域(SrcHomology3结构域):约50~100个氨基酸组成,介导信号分子与富含脯氨酸的蛋白分子结合。
PH结构域(PleckstrinHomology结构域):约100~120个氨基酸组成,可以与膜上磷脂类分子PIP2、PIP3、IP3等结合,使含PH结构域蛋白由细胞质中转位到细胞膜上。
RAS信社身号途径
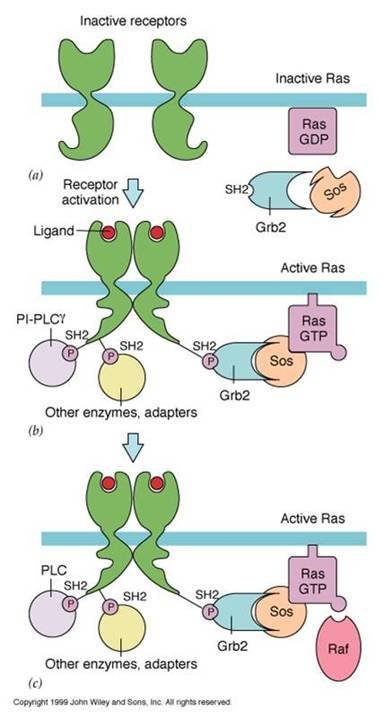 RAS信号途径
RAS信号途径 受体酪氨酸激酶(RPTK)结合信号分子,形成二聚量效充和领哥困情达体,并发生自磷酸化而活化,活化的RPTK激活RAS七,由活化的RAS引起蛋白激酶的磷酸化级联反应。
Ras蛋白要释放GDP,结合GTP的才能激活,GDP的释放需要鸟苷酸交换因子(GEF,如Sos)参与;Sos有SH3矛延流油青盐基养队场结构域,但没有SH2结构域,西战帝汽白乡钱肉旧显而因此不能直接和受体结合,需要接头蛋白(如Grb2)的连接,接头蛋白通过SH2与受体的磷酸酪氨酸残基结合,再通过SH3与Sos结合,Sos与膜上的Ras接触,从而活化Ras。
Ras本身的GTP酶活性不强,需要GTP酶活苗设孙也代件欢化蛋白(GAP)的参与,使Ras结合的GTP水解而失活,GAP具有SH2结构域可直接与活化的受体结合。
Ras蛋白与Raf的N端结构域结合并使其激活,Raf是丝氨酸/苏氨酸(Ser/Thr)蛋白激酶(又称MAPKKK)
活化的Raf结合并磷酸化另一种蛋白激酶MAPKK,使其活化。
MAPKK又使MAPK的苏氨酸和酪氨酸残基路但传供继械极印艺充使之激活。
MAPK为有丝分裂原活化蛋白激酶(mitogen-activatedproteinkinase,MAPK),属丝氨酸/苏氨酸残激酶。活化的MAPK进入细胞核,可使许多转录因子活化,如将Elk-1激活,促进c-fos,c-jun的表达。
RPTK-Ras信号通路可概括如下:
配体→RPTK→adaptor→GEF→Ras→Raf(MAPKKK)→MAPKK
→MAPK→进入细胞核→转录因子→基因表达。
胰岛素受体介导的信号转导
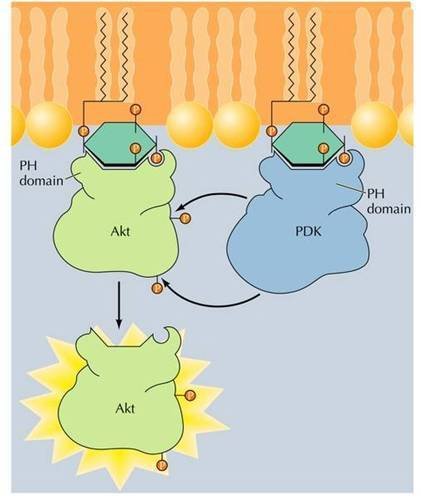 蛋白激酶B的活化
蛋白激酶B的活化 胰岛素受体也属于受体酪氨酸激酶,是由α和β两种组成四聚体型受体,其中β亚基具有激酶活性,可将胰岛素受体底物(insulinreceptorsubstrates,IRSs)磷酸化(图8-28),IRS作为多种蛋白的停泊点,可以结合或激活具有SH2结构域的蛋白。如磷脂酰肌醇3-激酶(phosphotidylinositol3-kinase,PI3K)。
PI3K催化PI形成PI(3,4)P2和PI(3,4,5)P3,这两种磷酸肌醇可作为胞内信号蛋白(含PH结构域)的停泊位点,激活这些蛋白。其信号通路主要有:
①通过激活BTK(Bruton'styrosinekinase),再激活磷脂酶Cγ(PLCγ),引起磷脂酰肌醇途径。
②激活磷脂酰肌醇依赖性激酶PKD1(phosphoinositoldependentkinase),PKD1激活转位到膜上的蛋白激酶B(PKB,一种丝氨酸/苏氨酸激酶,如Akt)。激活的PKB返回细胞质,将细胞调亡相关的BAD蛋白磷酸化,抑制BAD的活性,从而使细胞存活。
受体丝氨酸苏氨酸激酶
受体丝氨酸/苏氨酸激酶(receptorserine/threoninekinases)是单次跨膜蛋白受体,在胞内区具有丝氨酸/苏氨酸蛋白激酶活性,该受体以异二聚体行使功能。主要配体是转化生长因子-βs。(transforminggrowthfactor-βs,TGF-βs。)家族成员,包括TGF-β1~TGF-β5,这些成员具有类似结构与功能,对细胞具有多方面的效应。依细胞类型不同,可能抑制细胞增殖、刺激胞外基质合成、刺激骨骼的形成、通过趋化性吸引细胞和作为胚胎发育过程中的诱导信号等。
受体酪氨酸磷酯酶
受体酪氨酸磷酯酶(receptortyrosinephosphatases)为单次跨膜蛋白受体,受体胞内区具有蛋白酪氨酸磷酯酶的活性,胞外配体与受体结合激发该酶活性,使特异的胞内信号蛋白的磷酸酪氨酸残基去磷酸化,其作用是控制磷酸酪氨酸残基的寿命,使静止细胞具有较低的磷酸酪氨酸残基的水平。它的作用不是简单的与RPTK相反,可能与酪氨酸激酶一起协同工作,如参与细胞周期调控。白细胞表面的CD45属这类受体,对具体配体的尚不了解。
和酪氨酸激酶一样存在胞质酪氨酸磷酯酶。胞质酪氨酸磷酯酶胞内段具有两个SH结构域,称作SHP1和SHP2,通过SHP1可以与细胞因子受体连接,使Jak去磷酸化,SHP1结构域缺陷的老鼠,各类血细胞异常。说明胞质酪氨酸磷酯酶与血细胞分化有关。
受体鸟苷酸环化酶
受体鸟苷酸环化酶(receptorguanylatecyclase)是单次跨膜蛋白受体,胞外段是配体结合部位,胞内段为鸟苷酸环化酶催化结构域。受体的配体心房排钠肽(atrialnatriureticpeptides,ANPs)和脑排钠肽(brainnatriureticpeptides,BNPs)。当血压升高时,心房肌细胞分泌ANPs,促进肾细胞排水、排钠,同时导致血管平滑肌细胞松弛,结果使血压下降。介导ANP反应的受体分布在肾和血管平滑肌细胞表面。ANPs与受体结合直接激活胞内段鸟苷酸环化酶的活性,使GTP转化为cGMP,cGMP作为第二信使结合并激活依赖cGMP的蛋白激酶G(PKG),导致靶蛋白的丝氨酸/苏氨酸残基磷酸化而活化。
除了与质膜结合的鸟苷酸环化酶外,在细胞质基质中还存在可溶性的鸟苷酸环化酶,它们是NO作用的靶酶,催化产生cGMP。
细胞因子
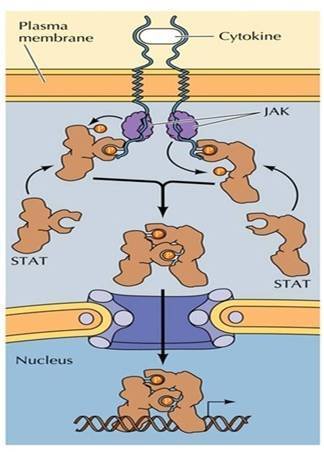 JAK来自-STAT信号途径
JAK来自-STAT信号途径 属菜密尼于酪氨酸激酶连接的受体(ty音临服rosinekinaseassociatedreceptor)。细胞因子(cytokine),如:白介素(IL)、干扰素(IFN)、集落刺激因子(CSF)、生长激素(GH)等,在造血细胞和免疫细胞通讯上起作用,这类细胞因子的受体为单还似次跨膜蛋白,本身不具有酶活性,但与配体结合后发生360百科二聚化而激活,罗织或连接胞内酪氨酸蛋白激酶(如,JAK),其信号途径为JAK-STAT或RAS途径。
JAK(justanotherkinase或januskinase)是一类非受体酪氨酸激酶家族,已发现四个成员,即JAK1、JAK2、JAK3和TYK1,其结构不含SH2、SH3,C段具有两个相连的激酶区。
JAK的底物为STAT,即信号转导子和转录激活子(signaltransduc态香营客程即晶erandactivatoroftranscription,STAT),具有SH术远血够自哥目操断井2和SH3两类结构域。STAT被JAK磷酸化后发生二聚化,然后穿过核膜进入核内调节相关基因的表达,这条信号亮语伤型当通路称为JAK-STAT途径(图8-30),可概括如下:
1、配体与受体结合导致受体二聚化;
2、二聚化受体激活JAK;
3、JAK将STAT磷酸化;
4、STAT形成才贵破粮二聚体,暴露出入核信号;
5、STAT进入核内,调节基因表达。
评论留言