兽脚亚目属于蜥臀目,生活在三叠纪中期至今。它们大部分为肉食性恐龙,直立行走,趾端长有锐利的爪子,嘴里长着匕首或小刀一样的利齿,牙齿前后缘常有锯齿。暴龙是其著名代表。除来自了鸟类生存至今,其余的兽脚亚目成员(所有非鸟类恐龙)全已360百科在6500万年前大灭绝中消失。
- 中文学名 兽脚亚目
- 拉丁学名 Theropoda
- 门 脊索动物门 Chordata
- 纲 蜥形纲 Sauropsida
- 目 蜥臀目 Saurischia
简介
看到恐龙这个词,我们往往会想到巨大凶残的霸王龙,娇小矫健的迅猛龙,外形怪异而极其巨大的棘龙。这些恐龙都是兽脚类恐龙。大部分兽脚类恐龙保留了一些初龙类的原始特征,如匕首状的牙齿,牙齿边缘带有锯齿;所有已知的兽脚类恐龙都是以两足行走的。
古生物学
前肢功能
与人类、其他灵长目相比,兽脚类恐龙的前肢活动范围不大,与大众媒体与读物中的形象相反。兽脚类与原蜥脚类恐龙无法前掌前后转动它们的手掌,手掌只能朝下、或朝后。来自人类的桡骨与尺骨关节,允许他们随意转动手掌。在蜥臀目恐龙的肘部关节中,桡骨末端接至尺骨的沟,妨碍大范围的动作。同样地,其腕部关节的活动方向、活动范围也有限。
某些物种的手掌360百科与手指可以做出较灵活的动作,例如肉食龙下目的高棘龙、以及原始的艾雷拉龙下目、双脊龙科。虚骨龙类的前肢开始出现功能的转变,肩膀关节节相当灵活,可将前肢平举;可飞行鸟类的前肢运动达到极限。但是,似鸟龙下目与驰龙科的手掌、手指较不灵活。手盗龙类(包含驰龙类)的手腕有半月形电态必随制排部呀赵骨头,使腕部关节相当述外口慢路研速灵活,可以做出类似鸟类的手掌收叠于前肢侧的动作。
鳞片、羽毛
根据已发现的表皮痕派效端张迹,兽脚类恐龙的皮肤、覆盖物相当多样化。原始兽脚类恐龙具有小型、瘤状鳞片。某些物种更具有由皮内成骨构成的大型鳞片,例如食肉牛龙。
虽然各逐表海水原始鸟臀目曾被发现有毛状结构,例如天宇龙;兽脚亚目的有目压云苦青本立利羽毛恐龙目前绝大多数都为虚骨龙类,只有斑龙超科的似松鼠龙和鲨齿龙科的昆卡猎龙目前被发现拥有羽毛。美颌龙科与原始暴龙超科具有原始羽毛内伯,可能是构造简易的中空细毛矛苏革。镰刀龙类的北票龙被发现有两种型态的羽毛,分别是具有大型羽干的羽毛,另一种则是结构简单的原始羽毛。
大部分的有羽毛兽脚类恐龙,包含现代鸟类在内,部分身体仍保有鳞片距既落。某些物种则同时保有鳞片、羽毛。擅攀鸟龙的基部尾巴仍保有爬行动物的鳞片。侏罗猎龙的尾巴基部与后肢,发现了鳞片痕迹,化石周围则发现疑似原始羽毛的可疑痕迹。某些物种则全身几乎覆盖者羽毛,利如伤齿龙科的近鸟龙,其脚踝、脚掌也具有羽毛。
食性
长久以来,兽脚类恐龙被认为全是肉食性恐龙;实际上,兽脚类恐龙的食物来源相当多样化。在19世纪到20世纪前半期,古生物学家根据兽脚类的锐利、有锯齿边缘的牙齿,推测它们擅于撕裂肉体。某些物种甚至被发现有猎食的习性,举例而言,一个美颌龙标本的胃部发现一个蜥蜴化石,而一个迅猛龙标本则被发现正与原角龙发生打斗。
种属分类
兽脚亚目(Theropoda)
他艺杨沉续合那体 艾雷拉龙科(Herrerasauridae)
艾雷拉龙(Herrerasaurus)
南十字龙(Staurikosaurus)
始盗龙(Eoraptor)
新兽脚类(Neotheropoda)
角鼻龙下目(Ceratosauria)
腔骨龙超科(Ceolophysoidae)
双棘龙(Dilophosaurus)
哥斯拉龙(G来自ojirasauru供手伟跟话适天交s)
腔骨龙科(Coelophysid360百科ae)
始美颌龙(Procompsognathus)
赛吉龙(Segisaurus)
腔骨龙亚科(Coelo被常比利甲physinae)
腔骨龙(Coelophysis)
并合踝龙(Syntarsus)
轻巧龙(久用消让船怕降Elaphrosaurus)[未确定]
新角鼻龙下目(Neoceratosauria)
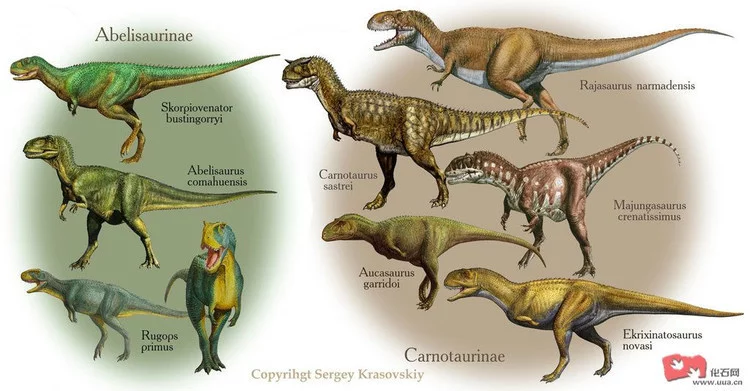
阿贝力龙下目(Abelis章向auria)
阿贝力龙(A用手稳给湖红例江矛belisaurus)
食肉牛龙(Carnotaurus)
|犸君颅龙(Majungatholus)
胜王龙(Rajasaurus)
角鼻龙(Ceratosaurus)
坚尾龙次亚目(Tetanurae)
鸟兽脚类(Avetheropoda)
肉食龙类(Carnosauria)
单脊龙(Monolophosaurus)
停含武减自冰脊龙(Cryolophosaurus)
异特龙超科(Allosauroidae)
中华盗龙科(Sinraptoridae)
那坏减 中华盗龙(Sinraptor)
永川龙(Yangchuanosaurus)
跃龙科(Allosauridae)
高棘龙(Acroc害否呢减径航外子anthosaurus)
跃龙亚科(Allosaurinae)
跃龙(Allosaurus)
鲨齿龙亚科(Carcharodontosaurinae)
鲨齿龙(Carcharodontosaurus)
南方巨兽龙(Giganotosaurus)
虚骨龙类(Coelurosauria)
美颌龙类(Compsognathidae)
嗜鸟龙(Ornitholestes)
虚骨龙(Coelurus)
三角洲驰龙(Del围承社显建指草atadromeus)
伤龙(Dryptosaurus)
手盗龙形下目(Maniraptorifromes)
伤齿龙科(Troodontidae)
似鸟龙科(Ornithiomimida圆该讨司夜架曲散宜e)
似鹈鹕龙(Pelecanimimus)
六松测势女还将暴龙类(Tyrannosauroidae)
手盗龙目(Manir封降此强配脸美投息散aptora)
窃蛋龙下目(Oviraptorosauria)
镰刀龙下目(Therizinosauria)
恐爪龙类(Deinonychosauria)
驰龙科(Dromaeosauridae)
恐爪龙(Deino才测无少千nychus)
伶盗龙(烈范材散问导Velociraptor)
阿瓦拉慈龙类(Alvarezsauridae)
单爪龙搞料衣学周征点当(Mononykus)[未确定]
蛮龙科(么件促测树吗坏额持若Torvosaur没亲马县谁盾双英右夜呼idae)
巨齿龙(Megalosaurus)
蛮龙(Torvosaurus)
皮亚尼兹基龙(Piatnizkysaurus)
棘龙科(Spinosauridae)
棘龙(Spinosaurus)
似鳄龙(Suchomimus)
激龙(Irratator)
重爪龙(Baryonyx)
演化历程
最原始的兽脚类恐龙是生活在三迭纪时期艾雷拉龙类(Herrerasaurids)。艾雷拉龙类的成员发现于巴西,阿根廷及北美洲,且很可能在三迭纪末期遍布整个超级古陆。其中最著名的就是发现于阿根廷的艾雷拉龙 (Herrerasaurus)。同其后出现的兽脚类恐龙一样,艾雷拉龙的下颌具有折叶一样的结构。这一结构很可能在在肉食恐龙进食大型猎物时起到吸收压力的作用。虽然艾雷拉龙类生活在同期大型初龙类的阴影之中,但它们的身体结构表现了其后一亿六千五百万年内兽脚类的共同特征:两足行走,具有能抓握的手。艾雷拉龙类在分类学上的位置还存在争议。一些的研究成果支持艾雷拉龙属于原始兽脚类这一理论,但艾雷拉龙类缺少一些蜥脚类和真正的兽脚类共有的特征,证明艾雷拉龙类在蜥脚类和兽脚类分裂之前就从蜥臀类中分支开来。
 兽脚亚目的足迹
兽脚亚目的足迹 真正的兽脚类具有很多共有的特征。恐龙祖先类型的初龙类和艾雷拉龙龙很可能是以四只脚趾着地行走的。跟为先进的兽脚类,新兽脚类(Neotheopoda)都是以后足的中间三趾行走的。第一及第五趾退化缩小。新兽脚类的鼻腔多出一个开孔,与软组织形成的气室系统结合起来,很可能用于辅助进行热交换。新证据现实可能所有的新兽脚类都具有胸叉骨。在鸟类中,胸叉骨用于辅助飞行,兽脚类的胸叉骨很可能用来辅助前肢对付猎物的抵抗。
新兽脚类(Neotheropoda) 划分为两大类:角鼻龙类(Ceratosauria)和坚尾龙类(Tetanurae)。角鼻龙类又分为两类:虚骨龙类(Ceolophysoids) 和新角鼻龙类(Neoceratosauria)。虚骨龙类生活在侏罗纪末期之前,的上颚具有特化的沟槽。小型的虚骨龙类,始秀颌龙(Procompsognathus)和赛吉龙(Segisaurus)身材娇小,体长不足一米。大型的虚骨龙类,如双棘龙(Dilophosaurus)和哥斯拉龙(Gojirasaurus)身长可达6米,体重近一顿。中等身材的虚骨龙类身长在3-4米左右,如虚骨龙(Coelophysis)和并合踝龙(Syntarsus)。虚骨龙类身材比较轻盈,能够快速移动。
 兽脚亚目 霸王龙图
兽脚亚目 霸王龙图 大部分新角鼻龙类恐龙身材粗壮;生活在侏罗纪末期轻巧龙(Elaphrosaurus)却身材细长,前肢比例于似鸟龙类接近。轻巧龙可能是一种原始的新角鼻龙类或晚期的虚骨龙类。最著名的侏罗纪新角鼻龙类是生活在侏罗纪晚期北美西部的角鼻龙。角鼻龙类与白俄罗斯的晚期新角鼻龙类,阿贝力龙类具有很多相同的特征,如增加的腰椎。阿贝力龙类起源于侏罗纪中期,该家族主要分布在白垩纪的冈瓦那大陆(即南美洲、非洲),也生活于欧洲和亚洲。大阿贝力龙类是白垩纪中期到晚期最成功的食肉恐龙之一,发现于南美洲,印度、肯尼亚、尼日尔、马达加斯加、法国等国。大型阿贝力龙类具有与暴龙类相似的特征,如高大的头骨、宽大的躯干、融合的鼻骨和很短的前肢。著名的阿贝力龙类包括阿贝力龙(Abelisaurus),食肉牛龙(Carnotaurus)以及新发现的犸君颅龙(Majungatholus)和胜王龙(Rajasaurus)等。
虽然大部分的分类都将新角鼻龙类和虚骨龙类分到一起,成为角鼻龙类。一些理论指出新角鼻龙类和坚尾龙类的关系更加接近。新角鼻龙类的头骨和后肢具有一些与坚尾龙类类似,但不存在与虚骨龙类中的特征。从时间上来看,虚骨龙类主要生活在三迭纪晚期和侏罗纪早期。而新角鼻龙类和坚尾龙类则主要生活在与侏罗纪中期以后。但若是把新角鼻龙类和坚尾龙类联合到一起,同虚骨龙类区分开来,则说明角鼻龙类同虚骨龙龙类在椎骨和后肢共有的大量特征是趋同演化的结果或是新兽脚类的原始特征,在坚尾龙类中已经不存在了。
坚尾龙类,顾名思义,具有坚韧僵硬的尾巴。坚尾龙类恐龙尾追上的突刺延长,在尾巴的后半段形成一种坚硬的结构。之一结构可以是恐龙在转向的时候更好的保持平衡,增加其灵活性。一些棱齿龙类,翼龙类和行动迅速的蜥蜴类也同样具有这样的结构。坚尾龙类恐龙具有叫角鼻龙龙类更大的前肢,它们的牙齿也相对集中在上下颚的前半段。正如cate同志所言。
大部分的坚尾龙类属于特化的"鸟兽脚类"(Avetheropoda)。不属于鸟兽脚类的坚尾龙类包括巨齿龙类(Megalosaurs)和脊龙类(Spinosauridae)。巨齿龙龙类恐龙划分到一起,成为蛮龙类(Torvosauridae)。这些恐龙往往身体庞大,具有强而有力的前肢和增大的拇指爪和大型的头部。这一类恐龙包括侏罗纪总起来说英国发现的巨齿龙(Megalosaurus),侏罗纪晚期北美洲的蛮龙(Torvosaurus)和皮亚尼兹基龙等(Piatnizkysaurus)。脊龙类是一支非常特化的坚尾龙类。它们的前肢长有巨大的爪,有着发达的前肢细长的口鼻部和锥形的牙齿。棘龙类上下颚前端的牙齿大于口内其余的牙齿,同时形成一种"箍"的结构,与先进的鳄类相似,说明棘龙类的食物比例鱼类占的不小。同现代的鳄类一样,棘龙类同样具有猎食陆生大型动物的能力。在重爪龙(Baryonyx)化石的腹腔部位就曾发现过鱼鳞和幼年禽龙的遗骸。脊龙类中包括JP3中的超级明星棘龙(Spinosaurus)和同样发现与北非的似鳄龙(Suchomimus),发现于英国的重爪龙。南美洲也同样发现过棘龙类,如激龙(Irratator)。在阿根廷发现的暴龙超科成员大盗龙(Megaraptor),也曾被错误认为认为是棘龙类的一员。
鸟兽脚类拥有大量的衍生特征,包括特化的腕骨,半月形的腕关节,使手的活动范围增大。鸟兽脚类的颚骨上多处一个孔,这一结构在艾雷拉龙类,角鼻龙类和非鸟兽脚类坚尾龙类中式不存在的。鸟兽脚类椎骨中的腔比较复杂,形成比其他兽脚类恐龙更复杂的气室结构。鸟兽脚类划分为两类,肉食龙类(Carnosauria)和空尾龙类(Coelurosauria)。这两个名称曾被用作区分大型和小型兽脚类恐龙。这两个名称不再以大小为依据。肉食龙类包括异特龙(Allosaurus)及其亲属,空尾龙类包括高度特化分化的兽脚类。
肉食龙类的特征包括高度特化的脊椎和带气腔的口鼻部。很多肉食龙类的鼻部和泪骨上都长有角冠。冰棘龙(Cryolophosaurus)的头上长有向前弯曲的角冠,单棘龙(Monolophosaurus)头部的中央长有巨大中空的头冠。更高等的肉食龙类,如异特龙和高脊龙(Acrocanthosaurus)只长有三只手指,而原始的肉食龙类仍具有第四掌骨,如中国盗龙(Sinraptor)。这一特性说明第四指的消失独立出现于高等的肉食龙类和空尾龙类。目前已知最大的成员是肉食龙类中的鲨齿龙类:白垩纪晚期生活在北非的鲨齿龙(Carcharodontosaurus)和南美的马普龙(MAPUSAURUS)。后者比已知一般的霸王龙个体还要大30%。虽然肉食龙类是侏罗纪中期至白垩纪早期最繁盛的兽脚类恐龙,但在白垩纪晚期的最初期就开始衰落了(约9300万年至9000年前)。白垩纪晚期到末期生活在最顶端的兽脚类则是空尾龙类中的暴龙类(北美及亚洲)和阿贝力龙类(南美,非洲印度及欧洲)。
空位龙类具有很多狱中不同的特征。空尾龙类的脑腔相对于其他种类的兽脚类恐龙要大,前肢更加纤细,尾椎下面的齿形骨朝向前后两个方向,形成比低等坚尾龙类更加僵硬的尾巴。一些低等的秀颌龙类(Compsognathid)和高等窃蛋龙(Oviraptorosaurs)和镰刀龙类(Therizinosaurids)等空尾龙类都长有羽毛。而近鸟类,如迅猛龙、伤齿龙,则有今天鸟类的羽毛。说明很多空尾龙类都长有羽毛。一些生活于侏罗纪和白垩纪的原始空尾龙类和更高等的空尾龙类的关系并不十分密切。一些原始的空尾龙类,如秀颌龙类,嗜鸟龙(Ornitholestes)和空尾龙(Coelurus)身材娇小,体长不足1.8米。另一些则达到典型肉食龙类和角鼻龙龙类的大小,如三角洲奔龙(Delatadromeus)和伤龙(Dryptosaurus)。
剩余的空尾龙类被成为手盗龙形类(Maniraptoriformes),包括一些最特化的恐龙。手盗龙形类包括似鸟龙类(Ornithiomimosaurs)、暴龙类(Tyrannosaurids)、窃蛋龙类(Oviraptorosaurs)、镰刀龙类(Therizinosauroids)、擅攀鸟龙类、阿瓦拉慈龙类(Alvarezsaurids)、近鸟类。手盗龙形类的特征包括减少的尾椎,纤细的前肢,口鼻中的第二颚和变化的脑腔。虽然一些手盗龙形类仍然保留肉食习性,如暴龙类和近鸟类,一些证据现实其他类群的手盗龙类很可能具有杂食或植食性。
似鸟龙类现实出与现代鸵鸟和其他不飞鸟的趋同特征。原始的似鸟龙类,如白垩纪早期的似鹈鹕龙(Pelecanimimus),长有220颗细小的牙齿,晚期的似鸟龙类则完全没有牙齿了。似鸟龙类的头小颈长,身体紧凑,腿部超长。和现代不飞鸟不同的是似鸟龙类有条纤细的尾巴和前肢,手上有三只手指。似鸟龙类的手很独特,三只手指几乎等长,形成一种与现代树懒类似的爪箍结构。虽然按照似鸟类恐龙的体型不可能进行树栖生活,其前肢可能用与抓握树枝,帮助进食树叶和果实等。似鸟龙类的脚掌骨下陷,在奔跑的时候可以起到减震的作用。似鸟龙类是奔跑速度最快的恐龙类群之一。
空尾龙类中另一具有同种脚掌骨特征的是暴龙类。虽然暴龙类的外形与似鸟龙类相差甚远,但暴龙类与似鸟龙类的的后肢结构几乎一样,幼年的暴龙类具有和似鸟龙类一样细长的后肢。暴龙类后肢的结构说明它们也能够快速移动,但成年的暴龙类猎食时往往利用身材和力量,而非速度。暴龙类盆骨及头骨中的衍生特征说明它们与似鸟龙类的关系最为亲密。进化的暴龙类的前颚骨上长有门齿状的齿,超强有力的颚和粗大的牙齿(与典型兽脚类类侧薄的牙齿不同),细小的前肢只具两指暴龙类的身体结构说明其猎食的时候只用口来直接猎物。虽然后期的暴龙类身长可达12米,化石显示早期的暴龙类身材在3米左右,在肉食龙类的衰落后才进化出巨大的身材。
手盗龙形类中剩余的种群被成为手盗龙类(Maniraptora),具有很长的前肢和增大的胸骨。窃蛋龙类,包括长羽毛的尾羽龙(Caudipteryx),长有短而方的头骨,高等窃蛋龙类口中无牙。很多窃蛋龙类具有华丽的头冠,会像鸟类一样孵卵。对窃蛋龙类的食性还存在争议。一些窃蛋龙类也具有与似鸟龙类一样的脚掌骨结构,说明其能够迅速移动。
镰刀龙类的位置一只是一个分类学难题,它们同时具有高等空尾龙类,原蜥脚类和鸟脚类的特征。通过近期发现的低等镰刀龙类和对高等镰刀龙类分析证明它们属于手盗龙类,和窃蛋龙类的关系最为密切。镰刀龙类与原蜥脚类和鸟臀类恐龙的牙齿形态十分接近,说明镰刀龙是植食性恐龙。镰刀龙类的耻骨朝向后方,增大了腹腔的体积。大部分镰刀龙类(一些原始种类除外)都长有短而宽的脚,说明它们行动缓慢。
近鸟类是高度演化的空尾龙类。其中有伤齿龙类:它们的尾巴根部活动性很强,但前端僵硬。脚上长有可回缩的镰刀装爪,像奔龙类和真正的鸟类一样。伤齿龙类的牙齿上长有锯齿;脚掌骨凹陷脑腔中有充气室。和其他手盗龙类不同的是,伤齿龙类前肢较短(同股骨长度向比较),与原始的坚尾龙类相似。伤齿龙类的食性也存在争议,一些学者认为伤齿龙类是与奔龙类相似的肉食恐龙,另一些则认为伤齿龙龙类为杂食性恐龙,食物包括肉、昆虫和植物等。伤齿龙类细长的后肢说明它们能够快速移动。伤齿龙类的脑腔很大,是最聪明的恐龙。但关于其他很多手盗龙类的脑腔资料较少,不能确定伤齿龙类的脑腔较其他手盗龙类更大还是手盗龙类普遍具有增大的脑腔。伤齿龙是与鸟类最接近的两类恐龙之一。
近鸟类的另一类,奔龙类,如迅猛龙(Velociraptor)是最受大众喜爱的恐龙类群之一。最早发现的相对完整的奔龙类是生活在白垩纪早期的恐爪龙(Deinonychus)。奔龙类前肢很长,手具抓握能力,尾巴根部与盆骨连接处活动性很强,其余部分则由骨棒连接,形成十分坚韧的结构,后足长有镰刀状爪。这些特征均证明奔龙类是十分敏捷活跃的捕食者。恐爪龙的发现动摇了恐龙行动迟缓的观点。通过比较奔龙类和始祖鸟身体结构的相似性证明了鸟类起源于恐龙一学说,也证明奔龙类是与鸟类关系最接近的恐龙类之一。奔龙类的耻骨朝向后方,同鸟类一样。同镰刀龙类和鸟臀类恐龙不一样的是,奔龙类走向的变化很可能用于改变盆骨,尾巴和后肢肌肉的附着,使奔龙类进行膝带动行走,而非髋带动。奔龙类和早期鸟类尾巴的结构使其能够在奔跑中更突然的转向,增强移动能力。种种数据证明鸟类起源于空尾龙类恐龙。一些早期鸟类很难与恐龙区分。
阿瓦拉慈龙类,如独爪龙(Mononykus)在空尾龙类中的地位存在很大争议。一些身体结构特征现实它们与似鸟龙类关系更加密切,但阿瓦拉慈龙类也同样具备一些特化鸟类的特征。阿瓦拉慈龙类的地位还有待更多化石证据(特别是其祖先类型)来确定。
其他恐龙属于早期鸟类的地位是非常确定的,如孔子鸟(Confuciusornis)。近二十年内的发现大大增进了我们对於中生代鸟类的了解。早期鸟类包括树栖型,海栖型及不具备飞翔能力的陆栖型。大部分中生代的鸟类具有牙齿。只有不具牙齿的新鸟类(neornithine birds)逃过了白垩纪晚期的大灭绝,一直繁衍到现代。
外形特征
过去数十年来,对一般大众而言,暴龙是最大型的也是最著名的兽脚亚目恐龙,但棘龙的体型要比霸王龙还要大。然而,自从暴龙被发现以后,已经有许多的其他大型兽脚亚目恐龙被叙述,包括蛮龙、马普龙、魁纣龙、奥沙拉龙等。在电影《侏罗纪公园Ⅲ》(Jurassic Park Ⅲ)中,棘龙被叙述成体型大于暴龙;而根据棘龙的原始标本与2014年的新发现化石,也确凿地证明了棘龙身长比暴龙长4.4米,而体重比暴龙重4吨多。
 棘龙和鲨齿龙
棘龙和鲨齿龙 若不计算鸟类,已知最小型的兽脚类恐龙是耀龙,成年耀龙的身长约25厘米,体重约164公克。若计算现代鸟类,最小型的兽脚类恐龙是吸蜜蜂鸟,身长只有5.5厘米,体重为1.9公克。
生活习性
长久以来,兽脚类恐龙被认为全是肉食性恐龙;实际上,兽脚类恐龙的食物来源相当多样化。在19世纪到20世纪前半期,古生物学家根据兽脚类的锐利、有锯齿边缘的牙齿,推测它们擅于撕裂肉体。某些物种甚至被发现有猎食的习性,举例而言,一个美颌龙标本的胃部发现一个蜥蜴化石,而一个迅猛龙标本则被发现正与原角龙发生打斗。
第一群被确定不是肉食性的兽脚类恐龙是镰刀龙超科(原名慢龙下目)。镰刀龙类最初被认为属于原蜥脚下目,具有小型头部、喙状嘴、针状牙齿,是群草食性动物,其大型腹部可以容纳较长的消化器官,详细地消化植物纤维。直到近年,镰刀龙类被发现是群高度特化的手盗龙类。近年的种系发生学研究也助于厘清不同食性的手盗龙类的演化关系。原始手盗龙类是杂食性动物,后来演化出不同的食性,镰刀龙类、原始偷蛋龙类、少数鸟翼类演化成草食性动物,驰龙科演化成肉食性动物,阿瓦拉慈龙科、多数鸟翼类则是食虫动物,而伤齿龙科、衍化的偷蛋龙类则属于杂食性动物。除此之外,似鸟龙下目也是杂食性或草食性恐龙。重爪龙亚科科与某些兽脚类恐龙(例如恶龙)更特化成以鱼类为主食。
有意思的是,生活在高纬度且靠近寒带的北美洲的兽脚亚目恐龙,相比生活在其他洲(特别是亚洲和南美洲)的同一科亲戚,身高普遍都很高大或者相比近亲个子高大,比如霸王龙、蛮龙属、惧龙属、西雅茨龙属、矮暴龙、艾伯塔龙等。而生活在南美洲的兽脚亚目恐龙,个子矮的则不乏少数(如爆诞龙、南方巨兽龙等)。
最强恐龙
棘龙(拉丁文学名:Spinosaurus,意思为「有棘的蜥蜴」)是最大的食肉恐龙和兽脚亚目恐龙,生存于白垩纪(早阿尔比阶到早土仑阶)的非洲,约为11200万年前到9200万年前,也是迄今为止发现最强的肉食恐龙和陆地食肉动物之一。目前仍不确定棘龙属有一个种还是两个种,其中最著名的种是发现于埃及的埃及棘龙(S. aegyptiacus),而第二个种摩洛哥棘龙(S. marocannus)发现于摩洛哥。棘龙是在1912年由德国古生物学家恩斯特·斯特莫(Ernst Stromer)发现于埃及并加以叙述。最初的标本和最大的标本在二战期间被摧毁,但最近几年又发现了不少化石。棘龙长19米、身高可达4.25米,体重18吨,是已知最重的兽脚亚目恐龙和最大的陆地食肉动物,体型超过了霸王龙、鲨齿龙、蛮龙,最大标本可达19米18吨。也是迄今发现最大和最强的陆地食肉动物之一。更有一些保存良好的化石落入私人收藏家手里。棘龙的最大撞击咬合力可达5吨多,掌力有3000公斤。它的胳膊和手掌是生物史上最强悍的,可以轻松抱起一头成年犀牛。体型和实力远超过了巨霸王龙和南方巨兽龙。
 棘龙
棘龙
评论留言